Mitokondriell kryss-kommunikasjon: Utfordringer for reduksjonistiske modeller for cellulær kommunikasjon
Av Otangelo Grasso. Oversatt herfra.
Følgende tekst ble inspirert av denne videoen: Er vi faktisk kontrollert av mitokondrier? Mindblowing nye funn
Abstrakt
 Nyere funn innen cellulær biologi har omdefinert vår forståelse av mitokondrier, og fremhever kommunikasjonssystemene deres. Langt fra å være bare energiprodusenter, engasjerer mitokondriene seg i sofistikert signalering, påvirker cellulær reparasjon, immunmodulasjon og systemisk regulering. Disse prosessene involverer mekanismer som tunnel-nanorør, Mitokondrie-Avledede Vesikler og retrograds signalveier, som alle krever tett koordinerte komponenter. I hjertet av dette systemet ligger lagring, koding og avkoding av biokjemiske signaler, for eksempel reaktive oksygenarter og membranpotensialer. Disse signalene krysser cellulære strukturer, avkodet av transkripsjonsfaktorer, og kulminerer med adaptive responser. Den gjensidige avhengigheten av disse mekanismene - spennende mitokondrier, kjernen og cytoskeletale elementer - avgir samtidig funksjonalitet, utfordrende trinnvis naturalistiske forklaringer. Denne kompleksiteten reiser spørsmål om hvordan slike gjensidig avhengige systemer kunne ha dukket opp uten eksisterende koordinering, og tilbyr en overbevisende sak for revurdering av reduksjonistiske modeller for cellulær kommunikasjon.
Nyere funn innen cellulær biologi har omdefinert vår forståelse av mitokondrier, og fremhever kommunikasjonssystemene deres. Langt fra å være bare energiprodusenter, engasjerer mitokondriene seg i sofistikert signalering, påvirker cellulær reparasjon, immunmodulasjon og systemisk regulering. Disse prosessene involverer mekanismer som tunnel-nanorør, Mitokondrie-Avledede Vesikler og retrograds signalveier, som alle krever tett koordinerte komponenter. I hjertet av dette systemet ligger lagring, koding og avkoding av biokjemiske signaler, for eksempel reaktive oksygenarter og membranpotensialer. Disse signalene krysser cellulære strukturer, avkodet av transkripsjonsfaktorer, og kulminerer med adaptive responser. Den gjensidige avhengigheten av disse mekanismene - spennende mitokondrier, kjernen og cytoskeletale elementer - avgir samtidig funksjonalitet, utfordrende trinnvis naturalistiske forklaringer. Denne kompleksiteten reiser spørsmål om hvordan slike gjensidig avhengige systemer kunne ha dukket opp uten eksisterende koordinering, og tilbyr en overbevisende sak for revurdering av reduksjonistiske modeller for cellulær kommunikasjon.
---
Nyere fremskritt innen cellulær biologi har belyst systemene som mitokondrier driver med intercellulær kommunikasjon, og avslører deres sentrale rolle i koordinerende cellulære og systemiske funksjoner. Disse funnene gir en forståelse av mekanismene og veien som er involvert, og kaster lys over det sofistikerte samspillet mellom cellulære strukturer og signalnettverk.
Bilde 1. Mitokondrier
Mitokondrier etablerer forbindelser gjennom en rekke traséer, hvorav den ene inkluderer Tunnel-NanoRør (TNR). Disse aktinrike fremspringene danner fysiske broer mellom celler, noe som tillater direkte overføring av mitokondrier. Denne prosessen er avgjørende for å reparere celleskader og gjenopprette metabolsk likevekt, spesielt i vev under stress. TNT-mediert mitokondriell overføring styres ved å signalisere molekyler som Reaktive OoksygenArter (ROA) og spesifikke proteinmarkører, som koder for nødvendigheten og retningen av overføringen. ROA -nivåer fungerer som en kode, signaliserer cellulær stress og metabolsk dysfunksjon. Spesifikke proteinmarkører, inkludert Miro1 og kinesinrelaterte proteiner, fungerer som adressekoder for å målrette mitokondrier mot mottakerceller.
 Kodingen av informasjon begynner ved givercellen, der mitokondriell dysfunksjon genererer stresignaler. Disse signalene lagres i form av biokjemiske gradienter, for eksempel ROS -akkumulering eller endringer i mitokondriell membranpotensial. Den kodede informasjonen overføres deretter gjennom TNT -er, med ROS som fungerer som en messenger for å kommunisere overføringshastigheten. Proteinmarkører på mitokondriell overflate fungerer som dekodere, gjenkjent av reseptorer i mottakerceller, og sikrer riktig dokking og integrering av organellen. Avkoding innebærer å tolke ROS -nivåer og overflateproteinmønstre, som mottakercellen behandler for å vurdere den funksjonelle statusen til de innkommende mitokondriene. Når den er dekodet, uttrykkes denne informasjonen gjennom metabolsk rekalibrering, inkludert restaurering av ATP -produksjon og reaktivering av cellulære reparasjonsveier. Dette dynamiske systemet fremhever raffinementet av cellulær kommunikasjon, med TNT-mediert overføring som både en ledning og en regulator for intercellulær signalering.
Kodingen av informasjon begynner ved givercellen, der mitokondriell dysfunksjon genererer stresignaler. Disse signalene lagres i form av biokjemiske gradienter, for eksempel ROS -akkumulering eller endringer i mitokondriell membranpotensial. Den kodede informasjonen overføres deretter gjennom TNT -er, med ROS som fungerer som en messenger for å kommunisere overføringshastigheten. Proteinmarkører på mitokondriell overflate fungerer som dekodere, gjenkjent av reseptorer i mottakerceller, og sikrer riktig dokking og integrering av organellen. Avkoding innebærer å tolke ROS -nivåer og overflateproteinmønstre, som mottakercellen behandler for å vurdere den funksjonelle statusen til de innkommende mitokondriene. Når den er dekodet, uttrykkes denne informasjonen gjennom metabolsk rekalibrering, inkludert restaurering av ATP -produksjon og reaktivering av cellulære reparasjonsveier. Dette dynamiske systemet fremhever raffinementet av cellulær kommunikasjon, med TNT-mediert overføring som både en ledning og en regulator for intercellulær signalering.
En annen mekanisme involverer Mitokondrie-Avledede Vesikler (MAV), som er spesialiserte vesikulære strukturer som bærer bioaktive molekyler til naboceller. Disse vesiklene fungerer som molekylære kurére, påvirker prosesser som betennelse og immunmodulasjon. Sammensetningen av disse vesiklene - lipidprofiler og spesifikk proteinlast - koder presis informasjon om cellulære tilstander og metabolske behov. Ved levering avkoder mottakerceller disse signalene, og integrerer dem i sine eksisterende veier for å koordinere systemiske responser.
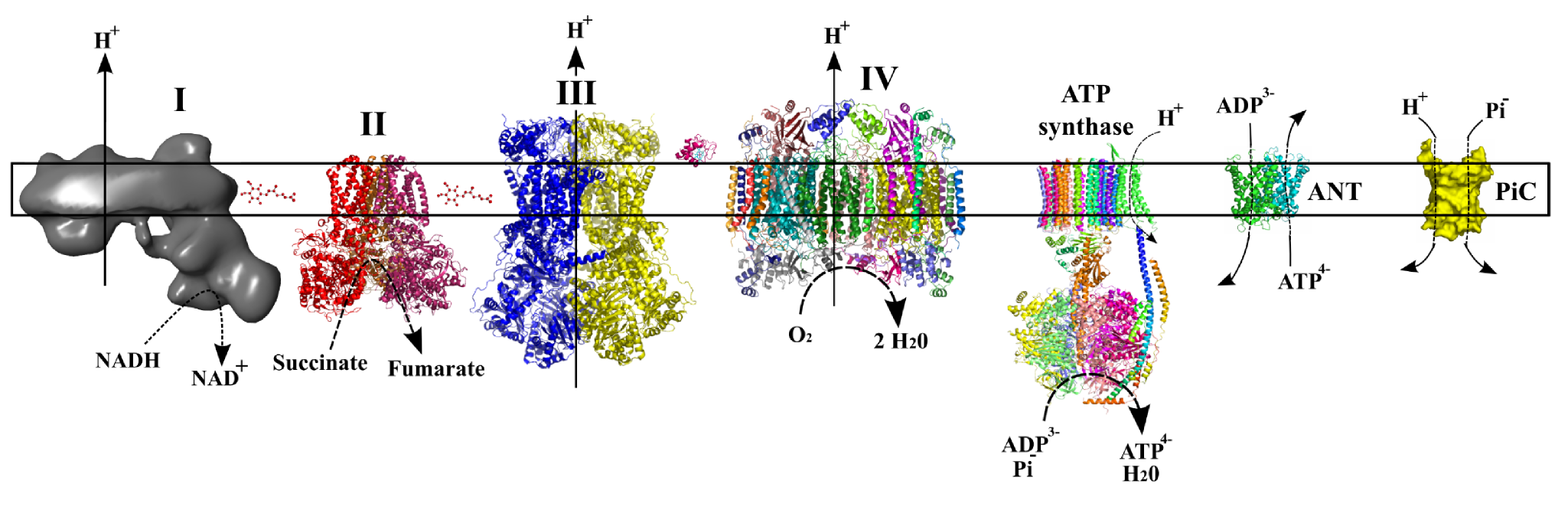
Bilde 2. Protonpumping gjennom mitokondrie
Den mitokondrielle utfoldede proteinresponsen (MUPR) eksemplifiserer en systemisk beskyttende mekanisme utløst av mitokondriell stress. Når proteiner i mitokondrier misfoldet, aktiverer MUPR kjerne-transkripsjonsprogrammer som oppregulerer molekylære chaperoner og proteaser. Denne toveis kommunikasjonen mellom mitokondrier og kjernen fremhever en sofistikert regulatorisk akse. Signaler som stammer fra mitokondriell dysfunksjon, forplanter seg til kjernen, noe som ber om genetiske responser som gjenoppretter proteostase og justerer metabolske aktiviteter med cellulære krav.
Kontaktsteder mellom mitokondrier og endoplasmatisk retikulum (ER), kjent som Mitokondrier-ER-KontaktSteder (Merks), letter utveksling av lipider https://snl.no/lipider og kalsiumioner. Disse interaksjonene er kritiske for å opprettholde energiproduksjon og lipidbiosyntese. Kalsiumioner fungerer for eksempel som budbringere i dette systemet, kjøreprosesser som ATP -generering og apoptose. Samspillet mellom disse organellene eksemplifiserer gjensidig avhengighet av cellulære rom for å opprettholde homeostase (likevekt).
 Mitokondriell kommunikasjon strekker seg til kjernen gjennom retrograd signalveier. Signaler fra mitokondrier, ofte initiert ved dysfunksjon, påvirker kjernens genuttrykk. Denne regulatoriske tilbakemeldingssløyfen er mediert av transkripsjonsfaktorer som NF-KB og p53, som styrer svar på betennelse og stress. Slike veier sikrer at mitokondriell funksjon er tett på linje med cellulære og systemiske krav, og understreker deres integrerende rolle i cellulær regulering.
Mitokondriell kommunikasjon strekker seg til kjernen gjennom retrograd signalveier. Signaler fra mitokondrier, ofte initiert ved dysfunksjon, påvirker kjernens genuttrykk. Denne regulatoriske tilbakemeldingssløyfen er mediert av transkripsjonsfaktorer som NF-KB og p53, som styrer svar på betennelse og stress. Slike veier sikrer at mitokondriell funksjon er tett på linje med cellulære og systemiske krav, og understreker deres integrerende rolle i cellulær regulering.
Bilde 3. Transportkjede gjennom mitokondrie
Mitochondriens innflytelse overskrider deres rolle i cellulær energiproduksjon. Mitokondrier inneholder unikt genetisk materiale og operativ autonomi. Denne autonomien beholder sin evne til å fungere som informasjonsprosessorer, og integrerer miljøsignaler for å drive cellulær tilpasning. Deres rolle strekker seg utover metabolsk regulering, påvirker nevral aktivitet, læring, hukommelse og til og med emosjonelle tilstander. Nyere studier avslører en toveis forbindelse mellom mitokondrier og psykologisk velvære, med emosjonelle responser målbare på mitokondrielt nivå, spesielt innen immunsystemceller.
 I tillegg til disse mekanismene, engasjerer mitokondriene seg i vesikulær transport for å lette metabolsk avfallshåndtering og lipidfordeling. Vesikler avledet fra mitokondrier er rettet mot lysosomer og peroksisomer, hvor de bidrar til autofagisk nedbrytning og resirkulering av ressurser. Denne prosessen er integrert i cellulær kvalitetskontroll, ettersom den eliminerer skadede organeller og omfordeler metabolske underlag for å opprettholde cellulær aktivitet.
I tillegg til disse mekanismene, engasjerer mitokondriene seg i vesikulær transport for å lette metabolsk avfallshåndtering og lipidfordeling. Vesikler avledet fra mitokondrier er rettet mot lysosomer og peroksisomer, hvor de bidrar til autofagisk nedbrytning og resirkulering av ressurser. Denne prosessen er integrert i cellulær kvalitetskontroll, ettersom den eliminerer skadede organeller og omfordeler metabolske underlag for å opprettholde cellulær aktivitet.
Integrerte eukaryote signalveier fremhever ytterligere sentraliteten i mitokondriell kommunikasjon i cellulær koordinering. Mekanismer som AMPK -signalering, som reagerer på cellulære energitilstander, og mTOR -veier, som regulerer mitokondriell biogenese, eksemplifiserer de komplekse interaksjonene som opprettholder cellulær funksjon. Disse traséene oversetter metabolske signaler til adaptive responser, og sikrer at energiproduksjon, ressursallokering og cellulær vekst er fint innstilt.
Bilde 4. Kinesin frakter en vesikkel-oppå mikrotubul
Den mitokondrielle krysskommunikasjonen illustrerer den sammenkoblede naturen til cellulær kommunikasjon. Ved å utnytte forskjellige mekanismer og veier, orkestrerer mitokondrier responser som er essensielle for å opprettholde cellulær og systemisk likevekt.
Systemisk gjensidig avhengighet av mitokondriell kommunikasjon
Mitokondrielle kommunikasjonssystemer eksemplifiserer gjensidig avhengighet, der flere cellulære aktører opererer på et samarbeidsvillig systemnivå vis. Denne gjensidige avhengigheten er avhengig av informasjonslagring, koding, overføring, avkoding og uttrykk, noe som nødvendiggjør samtidig eksistens av forskjellige komponenter for funksjonalitet.
Informasjon lagring og koding
 Mitokondrier lagrer informasjon gjennom biokjemiske gradienter, så som reaktive oksygenarter (ROA) nivåer og mitokondriell membranpotensial. Disse gradientene koder for stresssignaler, som videre er foredlet av spesifikke proteinmarkører, så som MiRO1 og kinesinrelaterte proteiner. Kodingsmekanismen er avhengig av mitokondriers kapasitet til å generere og opprettholde disse gradientene, og danner et grunnleggende lag for cellulær signalering.
Mitokondrier lagrer informasjon gjennom biokjemiske gradienter, så som reaktive oksygenarter (ROA) nivåer og mitokondriell membranpotensial. Disse gradientene koder for stresssignaler, som videre er foredlet av spesifikke proteinmarkører, så som MiRO1 og kinesinrelaterte proteiner. Kodingsmekanismen er avhengig av mitokondriers kapasitet til å generere og opprettholde disse gradientene, og danner et grunnleggende lag for cellulær signalering.
Overføringsmekanismer
Kodede signaler overføres gjennom Tunnel NanoRør (TNR) eller Mitokondrie-Avledede Vesikler (ADV). TNR-er letter direkte overføring av mitokondrier mellom celler, mens ADV-er innkapsler bioaktive molekyler, og leverer lasten til fjerne mål. Disse traséene krever presis koordinering av cytoskeletal dynamikk {mikrotubuler etc -oversetters tilføyelse} og vesikulær trafikkvirksomhet, og sikrer troskap i signallevering.
Bilde 5. Hver av de 20 aminosryrene har (også) sine koder. Kreditt: Otangelo Grasso
Avkodingsprosesser
Mottakerceller avkoder innkommende signaler ved bruk av reseptorproteiner og transkripsjonsfaktorer som tolker ROA -nivåer, membranpotensialer og vesikulært innhold. For eksempel formidler transkripsjonsfaktorer som NF-κB og p53 genetiske responser på mitokondriesignaler, og justerer cellulære aktiviteter med metabolske krav.
Uttrykk og systemisk innvirkning
Avkodede signaler manifesterer seg ved metabolsk rekalibrering, aktivering av reparasjonsveier eller cellulær differensiering. Denne uttrykksfasen integrerer mitokondriell inngang i systemiske prosesser, påvirker nevral aktivitet, immunrespons og energifordeling. Slik integrasjon demonstrerer gjensidig avhengighet av mitokondrielle signalnettverk og cellulære maskiner.
 Viktige gjensidig avhengighets spillere
Viktige gjensidig avhengighets spillere
For at dette systemet skal operere, må flere komponenter dukke opp samtidig:
- Mitokondrielle ROA -produksjons- og reguleringssystemer
- Proteinmarkører for signalkoding og målretting
- Cytoskeletale strukturer som muliggjør TNR
- Vesikkel -lenke; dannelse og transportvirksomhet
- Cellulære reseptorer og transkripsjonsfaktorer for avkoding
- Nukleært genetisk maskineri for respons-uttrykk
Bilde 6. Organisering av kompleksutet -et kjennetegn ved livet
Disse elementene må eksistere samtidig for å muliggjøre funksjonell kommunikasjon. Fraværet av noen komponent forstyrrer hele systemet, og fremhever usannsynligheten av gradvis, trinnvis montering.
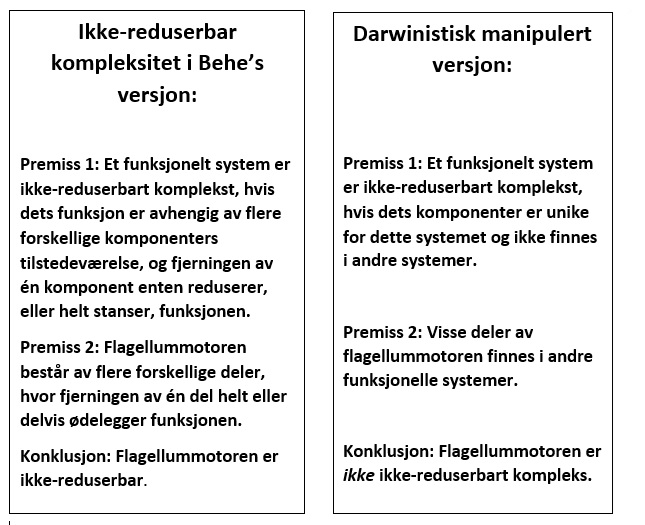 Åpne spørsmål og konseptuelle utfordringer
Åpne spørsmål og konseptuelle utfordringer
1. Samtidig fremvekst av komponenter
Hvordan kan alle nødvendige komponenter for mitokondriell kommunikasjon dukke opp samtidig? Hvert element er avhengig av de andre for funksjon, og lager et system som trosser gradvis montering.
Konseptuelt problem: ikke-reduserbar gjensidig avhengighet
- Systemer som TNR og MDA-er krever eksisterende cytoskeletale og vesikulære maskiner.
- Fraværet av en komponent gjør systemet ikke-funksjonelt.
Bilde 7. En krig om begreper
2. Opprinnelsen til kodede biokjemiske signaler
Hvilken mekanisme står for opprinnelsen til biokjemiske gradienter, for eksempel ROA og membranpotensialer, som pålitelige signalkoder?
Konseptuelt problem: Koding av presisjon
- ROA -nivåer krever tett regulering for å fungere som signaler uten å indusere cytotoksisitet.
- Generering og tolkning av disse gradientene nødvendiggjør kompleks eksisterende regulering.
3. Koordinering over cellulære rom
Hvordan oppsto mekanismer som muliggjorde kryssprat mellom mitokondrier, kjernen og andre organeller?
Konseptuelt problem: Koordinering av kommunikasjon på kryss av avdelinger.
- MERK-er krever synkroniserte lipid- og kalsium-utvekslingsmekanismer.
MERK-er krever synkroniserte lipid- og kalsium-utvekslingsmekanismer.
- Retrograd signalering krever presis interaksjon mellom mitokondrielle og kjerne systemer.
Bilde 8. Hvem av dem kom først -sirkulær avhengighet, Kreditt: Otangelo Grasso
4. Evolusjon av transkripsjonsregulering
Hva forklarer opprinnelsen til transkripsjonsfaktorer som NF-κB og p53 som medierer mitokondrielle signaler?
Konseptuelt problem: Regulatorisk kompleksitet
- Disse faktorene må forhåndseksistere for å svare på mitokondrielle innganger, og skaper en sirkulær avhengighet.
- Deres engasjement i bredere cellulære prosesser kompliserer forklaringer på deres fremvekst.
5. Systemnivåintegrasjon
Hvordan oppstod integrasjonen av mitokondrielle signaler i nevrale, immune og metabolske systemer?
Konseptuelt problem: Multisystem interavhengighet
- Nevrale og immunresponser på mitokondrielle signaler krever eksisterende systemiske rammer.
 - Sammenkoblingen til disse systemene motstår forklaring gjennom ikke-styrte prosesser.
- Sammenkoblingen til disse systemene motstår forklaring gjennom ikke-styrte prosesser.
Bilde 9. Genregulerings-nettverket kan sammenlignes med kretskort... Kreditt: Otangelo Grasso
Konklusjon: Kommunikasjons-systemene som involverer mitokondrier fremhever en gjensidig avhengighet mellom cellulære komponenter. Disse systemene krever samtidig funksjonalitet på tvers av forskjellige veier, og utfordrer forestillingen om gradvis, ikke-styrt evolusjonær fremvekst. å adressere disse åpne spørsmålene krever å utforske rammer som strekker seg utover naturalistiske forutsetninger, slik den samarbeidsvillige kompleksiteten som er observert i mitokondriell kommunikasjon, trosser reduksjonistiske forklaringer.
Oversatt, via google oversetter, og bilder ved Asbjørn E. Lund